Het immuunsysteem en de immuniteit bij varkens (deel 2)

Een specifieke humorale immuunrespons begint wanneer naïeve B-lymfocyten hun specifieke antigeen in de secundaire lymfoïde organen en weefsels herkennen (zie pathogeenherkenning).
Elke B-lymfocyt heeft een antigeenreceptor (BCR: B-cel receptor), een oppervlakte-immunoglobuline (IgM of IgD), die zich bindt aan specifieke antigeendomeinen die antigeendeterminanten of epitopen worden genoemd.
De specificiteit van een antigeenbindingsplaats hangt af van de genen die coderen voor variabele domeinen (VH en VL); bij zoogdieren kan een enorm aantal genen en, bijgevolg, verschillende BCR worden gegenereerd door genrecombinatie en / of genconversie, waardoor een dier in staat wordt gesteld om bijna alle vreemde antigenen te herkennen.
Na herkenning wordt elke specifieke B-lymfocyt geactiveerd, ondergaat het klonale proliferatie en differentieert het tot een plasmacel (of antilichaamsecreterende cel, ASC) die in staat is om immunoglobulinen (antilichamen), elk specifiek voor zijn epitoop, te synthetiseren en uit te scheiden.
De specifieke humorale immuunrespons is geoptimaliseerd dankzij de hulp van T-cellen.
Aanvankelijk scheidt de B-cel IgM af in een monomere vorm en met identieke antigene specificiteit.
Met betrekking tot het type antilichaamsecretie worden antigenen geclassificeerd als T-celafhankelijk of T-celonafhankelijk, afhankelijk van het feit of T-celhulp nodig is om een antilichaamrespons te beginnen.
T-celonafhankelijke antigenen (bijvoorbeeld bacterieel lipopolysaccharide, flagelline, pneumococcus-polysaccharide, enz.) kunnen alleen IgM-antilichamen met een bescheiden affiniteit en zonder B-celgeheugen induceren.
Vrijwel alle vreemde antigenen zijn T-cel-afhankelijke antigenen en vereisen, voor een volledige activering van B-lymfocyten, een aanvullend signaal (cytokine) door specifieke T-helpercellen die hetzelfde antigeen herkennen (samenwerking tussen T en B-cellen).
In dit laatste geval werkt de B-cel als een antigeen presenterende cel die het bewerkte antigeen presenteert aan T-helpercellen en die verwante herkenning maakt Ig isotype-switching (antilichaamklasse-omschakeling) van IgM naar een verschillende klasse van immunoglobulinen, IgA, IgE of IgG mogelijk, afhankelijk van het uitgescheiden Th-cytokine en betrokkenheid bij B-celstimulatie (respectievelijk IL-4 of IFNγ). Een ander cruciaal signaal voor het omschakelen van antilichamen is de betrokkenheid van de CD40-receptor op B-cellen door CD154 (CD40L).
Onder deze "hulp" ondergaan prolifererende B-cellen somatische hypermutatie van oppervlakte-immunoglobulinen en zullen dan worden voorzien van een specifiek isotype. Deze B-lymfocyten worden verder geselecteerd op basis van een optimale Ig-affiniteit voor het antigeen door in nauw contact te komen met de folliculaire dendritische cellen die hen het antigeen zelf presenteren. Alleen B-lymfocyten die een hoge antilichaamaffiniteit voor het antigeen bezitten en deze kunnen verwerken en presenteren, zullen positief worden geselecteerd. Aldus overleven alleen deze laatste B-lymfocyten, prolifereren en differentiëren tot plasmacellen die antilichamen van een specifieke isotype-klasse synthetiseren en uitscheiden, met een enkele specificiteit en hoge affiniteit, waardoor het vermogen om het antigeen te binden wordt verbeterd en derhalve de pathogenen worden geneutraliseerd en vernietigd.
Wanneer de primaire immuunrespons eindigt, zullen sommige van deze B-cellen en plasmacellen die tijdens dit eerste contact zijn geprimed en geselecteerd, overleven als geheugencellen met een lange levensduur, klaar om te worden geactiveerd bij daaropvolgende blootstelling aan hetzelfde antigeen (secundaire immuunrespons) waarbij dan dezelfde antilichaamklasse maar met een hogere titer en verhoogde affiniteit geproduceerd wordt.
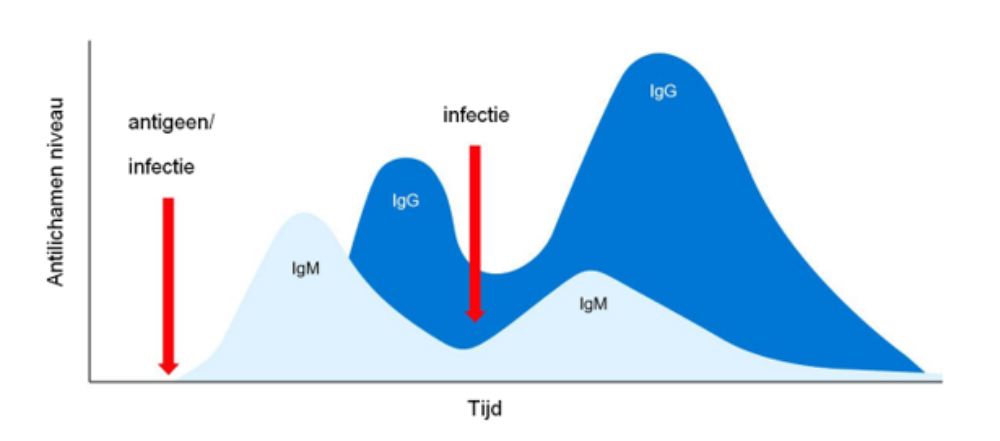
Samenvattend, in de primaire humorale immuunrespons wordt eerst een enkele klasse IgM in monomere vorm en met identieke antigene specificiteit uitgescheiden, vervolgens, na enkele dagen, als gevolg van isotype switching, treedt een secretie van een andere Ig-klasse, bijvoorbeeld IgG op. De secundaire humorale immuunrespons, geïnduceerd door een daaropvolgende blootstelling aan hetzelfde antigeen, zal voornamelijk gekenmerkt worden door de productie van IgG-isotype maar met een hogere titer en affiniteit (Figuur 1).
Antistoffen kunnen optreden tegen pathogenen met behulp van verschillende verdedigingsmechanismen:
- Remming van infectiviteit van pathogenen door directe binding daaraan (neutralisatie, attachment blocking, immunologische exclusie).
- Activering van alternatieve (opsonisatie) of klassieke complementroutes met pathogene lyse.
- Bij het coaten van het pathogeen (of geïnfecteerde cellen), stellen antilichamen accessoire cellen, die de kristalliseerbare fragment (Fc) delen herkennen, in staat om het pathogeen op te nemen en te doden door opsonisatie en fagocytose of om geïnfecteerde cellen te doden door ADCC ("Antilichaam-afhankelijke cellulaire cytotoxiciteit" ).
- Regulering van immuunreacties door interactie met Fc-receptoren (FcR).
FcR’s worden tot expressie gebracht door vele aangeboren immuuneffectorcellen zoals mastcellen, neutrofielen, monocyten, macrofagen en dendritische cellen. In het bijzonder activeren de FcyR's (Fc-receptor voor IgG), bij binding aan immuuncomplexen, de activering van aangeboren cellen en hun effectorreacties (fagocytose, oxidatieve burst, cytotoxiciteit, afgifte van vasoactieve peptiden en chemo-attractans). FcR's op dendritische cellen bevorderen het vangen, verwerken en presenteren van antigeen in cytotoxische T- en T-helper subsets.
Omgekeerd is de remmende FcR voor IgG, die tot expressie wordt gebracht op B-cellen, betrokken bij de regulatie van B-celactivatie en -ontwikkeling en in plasmacel overleving.
Vertaald uit het Engels. Oorspronkelijk artikel van de Universiteit van Parma, afdeling immunologie, Italië, 6 november 2017.